The Ubiquitination of PINK1 Is Restricted To Its Mature 52-kDa Form
Parkinson’s disease (PD) is an adult-onset neurodegenerative disorder that is characterized histologically by the loss of ventral midbrain dopaminergic neurons and by the presence of intracytoplasmic proteinaceous inclusions, or Lewy bodies, in the few neurons that are spared (Dauer and Przedborski, 2003). In a subset of PD cases, mutations in PINK1, which encodes for a mitochondrial kinase, have been associated with an early-onset form of the disease, inherited as an autosomal recessive trait (Valente et al., 2004).
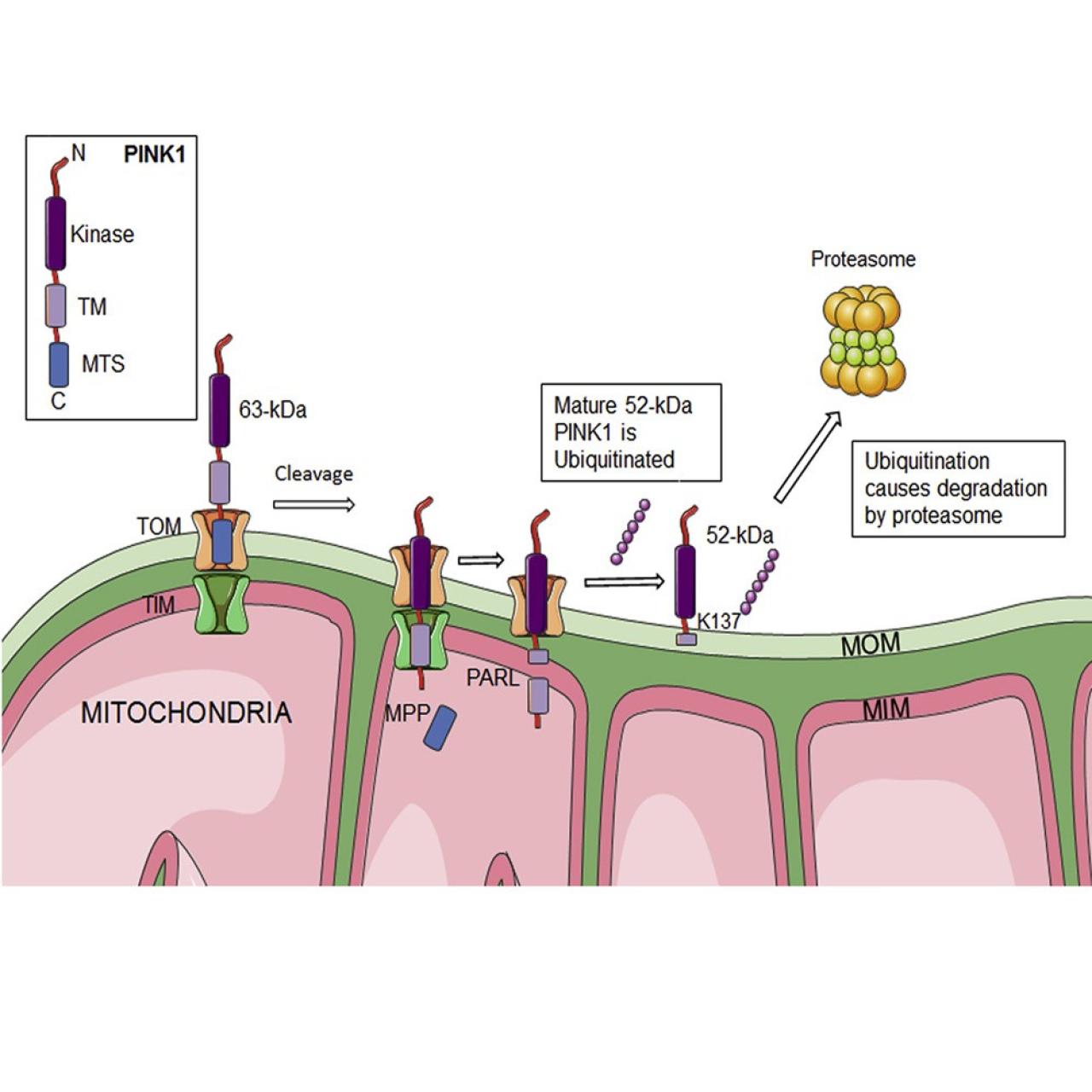
In energized mitochondria, full-length PINK1 (63 kDa) is rapidly imported and processed in two steps: first by the mitochondrial processing peptidase (MPP) to a 60-kDa intermediate form (cleavage between aa 34 and 35 [Greene et al., 2012; Jin et al., 2010]), and then by the presenilin-associated rhomboid-like protein (PARL) to a 52-kDa mature form (cleavage between aa 103 and 104 [Greene et al., 2012; Jin et al., 2010; Whitworth et al., 2008]). The 52-kDa form is located on the mitochondrial outer membrane (MOM) and retains most of the known functional properties of the full-length form (Sim et al., 2012).
Whereas the transcription of PINK1 is high (Blackinton et al., 2007; Taymans et al., 2006; Unoki and Nakamura, 2001; Zhou et al., 2008), the steady-state levels of all three PINK1 protein species are unexpectedly low (Zhou et al., 2008). Although low mRNA stability might account for this finding, a second possibility is that some of the PINK1 polypeptide species, and especially the fully processed mature 52-kDa form, are turned over rapidly. In support of this second possibility, it has been found that PINK1 is ubiquitinated, but which of the three PINK1 species is modified is not known (Shiba et al., 2009; Zhou et al., 2008).
We show here that mature, but not full-length, PINK1 is polyubiquitinated and that its level is kept low in healthy mammalian mitochondria via a specific ubiquitination at Lys-137 (K137), followed by subsequent proteasomal degradation.
Results
Ubiquitinated PINK1 Is Anchored Primarily to Mitochondria.
The ubiquitin/proteasome system is known to degrade MOM proteins, such as mitofusins (Tanaka et al., 2010), in damaged mitochondria. To test whether the same process applies to PINK1, we first asked whether ubiquitin is attached covalently to PINK1. We thus transfected influenza hemagglutinin (HA)-tagged ubiquitin (HA-Ub) into a stable human HEK293T cell line, in which exogenous PINK1 expression is ∼3–5 times less than that in transiently transfected cells in the presence or absence of the proteasome inhibitor MG132. As described previously (Vives-Bauza et al., 2010), lysates from transfected cells with normal mitochondrial membrane potential were immunoprecipitated with anti-PINK1 and western blotted to reveal the HA tag. A HA-immunopositive smear was detected, most prominently after treatment with MG132 (Figure 1A, upper panel). When these blots were re-probed with anti-PINK1, an immunopositive smear was observed (Figure 1A, lower panel), implying that PINK1 is ubiquitinated covalently. In addition, these polyubiquitinated PINK1 (Ub-PINK1) bands were resistant to treatment with Empigen BB, a denaturing detergent to weaken protein-protein interaction without disrupting the high-affinity antigen-antibody interactions (Figure S1A), supporting that polyubiquitinated bands were generated directly from PINK1. Ub-PINK1 was also observed in other cell lines treated similarly (Figures S1B and S1C). We also detected Ub-PINK1 in cells harboring endogenous PINK1, ubiquitin, or both (Figures S1D–S1G), implying that the endogenous ubiquitination machinery can modify endogenous PINK1. Of note, among different clonal cell lines that we screened, we only consistently detect endogenous Ub-PINK1 species (Figure S1G) in a BE(2)-M17 neuroblastoma cell line that stably expresses human wild-type α-synuclein (Guardia-Laguarta et al., 2014). Finally, upon exposure of cells to 1 μM valinomycin to collapse ΔΨm, Ub-PINK1 was reduced dramatically in experimental conditions described above (Figures 2F and S1H). These results indicate that Ub-PINK1 is a prominent feature of healthy mammalian mitochondria.